A phylogeny gives an evolutionary context to an ecological story
Here in the Cowman Lab, we use phylogenetics and bioinformatics to study the origin and diversification of tropical reef assemblages.
Our research focuses on three main areas:
Our research focuses on three main areas:
1. Resolving the Tree of Life on coral reefs
Molecular phylogenetics has become the fundamental tool used by the global community of evolutionary biologists to gradually teased apart the evolutionary relationship of what Darwin envisioned as the “tangled bank” of life on Earth. Aside from its utility in taxonomy and systematics, a phylogeny can provide a valuable framework in which we can study ecological patterns and processes while accounting for species ancestry. In the Cowman Lab, we are actively increasing the phylogenetic resolution for reef-associated fishes and coral lineages by leveraging publically available genetic data and generating new phylogenomic datasets.

2. Patterns of biodiversity & biogeography: past present and future
|
Despite the lack of physical geographic barriers across the Indo-Pacific gradient, discrete provinces can be outlined based on assemblage similarity. Geographic delineation is an important first step to establish and evaluate conservation priorities, but it can also provide a deeper understanding of the evolutionary forces shaping biodiversity. In the case of tropical reef assemblages, there are a number of schemes that delineated realms, regions, provinces and ecoregions based on shared environmental traits; composition of endemic taxa; and measures of species dissimilarity. But the challenge still remains to synthesise these assemblage level patterns, and the shared histories they represent, to disentangle their evolutionary cause. For instance, it remains unclear whether any provincial scheme based on current species distributions has had macroevolutionary significance in the past (Cowman, 2014). In the Cowman Lab, we are integrating phylogenetics, ancestral range estimation and biodiversity metrics to examine how patterns of biodiversity and provinciality have changed through time and how past climatic and geological changes have shaped present-day patterns of biodiversity across the marine tropics (see Cowman et al., 2017)
|
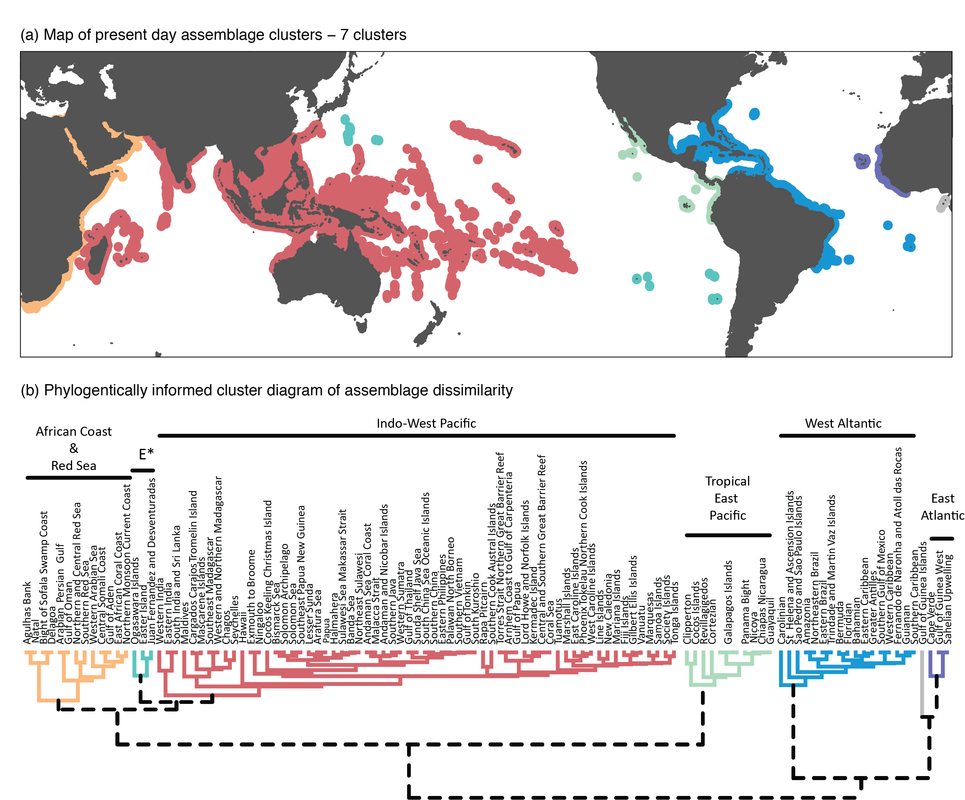
Figure from Cowman et al., (2017) - The biogeography of tropical reef fishes: Endemism and provinciality through time. Figure shows tropical provinciality of extant reef fish assemblages based on phylogenetic dissimilarity clustering methods. (A) Map of ecoregions; colour indicates their association with a particular cluster. (B) Dendrogram showing the clustering of ecoregions based on phylogenetic dissimilarity
|
3. The relationship between rate of molecular evolution, species richness and life history
An aspect of evolutionary biology with major implications for the study of biodiversity is how patterns of genomic change can influence, and be influenced by, macroevolutionary patterns. There is persistent evidence across the Tree of Life where clades with higher species richness tend to have faster rates of molecular evolution. But it is not clear if faster molecular rates are a cause, or consequence, of higher speciation. This relationship has yet to be explored for the longitudinal gradient in coral reef diversity. Specifically, do coral reefs in the IAA support faster rates of molecular evolution? The link between species richness and the molecular rate could be reflected across the Indo-Pacific biodiversity gradient in several ways. In the Cowman Lab, we are investigating how rates of molecular evolution in reef-associated fish and coral are linked to life history, range size and rates of diversification. We are developing new phylogenomic datasets to help us explore these questions.
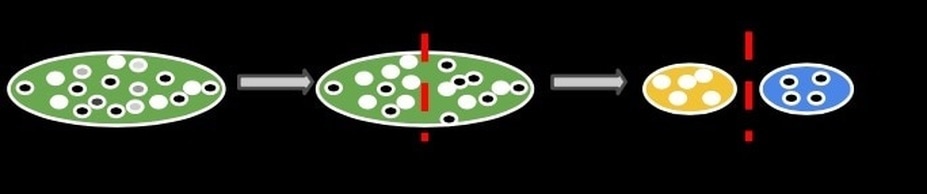
Do accelerated rates of molecular evolution increase the chance of speciation? Or does speciation result in accelerated rates of molecular evolution? The former implies that an organism with a higher rate of mutation across its genome will have a higher probability of speciation through the acceleration of hybrid incompatibilities. The latter implies that speciation is a special process, where incipient species generally have smaller population sizes, increases the change of mutations going to fixation, subsequently inflating observed rates of substitution.
